1. El ciclo celular.
El ciclo celular se divide en dos etapas: la interfase y la fase M o de división.
-Interfase: la célula realiza sus funciones vitales habituales y, en su caso, se prepara para la división. A su vez, consta de 3 fases sucesivas:

 La meiosis es un mecanismo de división celular especial por el que se forman gametos, es decir, células con la mitad de los cromosomas que la célula madre. La meiosis es exclusiva de los organismos con reproducción sexual
La meiosis es un mecanismo de división celular especial por el que se forman gametos, es decir, células con la mitad de los cromosomas que la célula madre. La meiosis es exclusiva de los organismos con reproducción sexual
El ciclo celular se divide en dos etapas: la interfase y la fase M o de división.
-Interfase: la célula realiza sus funciones vitales habituales y, en su caso, se prepara para la división. A su vez, consta de 3 fases sucesivas:
- Fase G1: etapa inmediatamente posterior a la mitosis. Etapa de crecimiento celular y de intensa síntesis de ARN y proteica, necesaria para el crecimiento.
- Fase S: cuando la célula alcanza un tamaño crítico necesita dividirse. Comienza así la síntesis de ADN y la duplicación de cromosomas, que quedan formados por dos cromátidas.
- Fase G2: la célula se prepara para la división. Termina su crecimiento y se sintetizan ARN y proteínas.
-Fase M: es la fase de división celular. Incluye la mitosis y la citocinesis.
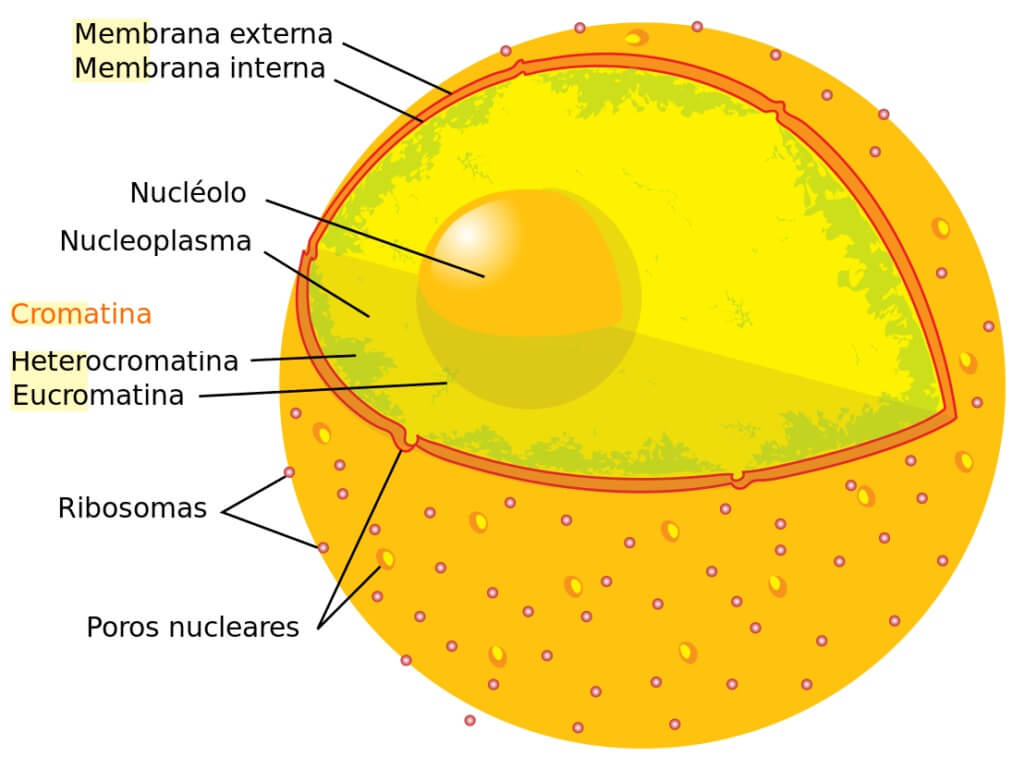
2. Núcleo interfásico.
El núcleo contiene el material genético en forma de ADN y dirige la actividad celular. El núcleo posee una envoltura con poros, un nucleoplasma o contenido nuclear con cromatina y, habitualmente, uno o más corpúsculos esféricos llamados nucleolo.
-Envoltura nuclear: es una doble membrana, con un espacio perinuclear.
- Membrana nuclear externa: es una continuación de la del RER y puede presentar ribosomas en su cara citosólica.
- Membrana nuclear interna: en su cara interna lleva una red de filamentos proteicos, la lámina fibrosa o lámina nuclear.
- Poros nucleares: es un orificio con una estructura proteica llamada complejo del poro.
-Nucleoplasma: medio interno similar al citosol. Contiene una disolución de sales, nucleótidos, ARN y proteínas.
-Nucleolo: estructura más o menos esférica, densa y de contorno irregular. Aquí se sintetizan los ARNr y se ensamblan para formar ribosomas.
-Cromatina: en el núcleo interfásico el ADN siempre va asociado a proteínas, formando la cromatina. La cromatina presenta diferentes niveles de complejidad:
- Nucleosoma y collar de perlas.
- Fibra de cromatina.
3. El núcleo mitótico: cromosomas.
El cromosoma mitótico es un cromosoma doble muy condensado y listo para poder repartir cada cromátida a una célula hija.El cromosoma metafásico consta de los siguientes elementos:
- Cromátida: cada parte simétrica y genéticamente idéntica.
- Constricción primaria o centrómero: estrechamiento que sirve de zona de unión de las dos cromátidas hermanas.
- Cinetocoros: complejos proteicos situados a ambos lados del centrómero. Son las zonas donde se unirán los microtúbulos cinetocóricos, que permitirán la separación de las cromátidas en la anafase.
- Constricciones secundarias: estrechamientos asociados a los nucleolos y llamados regiones de organización nucleolar (RON).
- Telómero: fragmentos terminales de ADN que protegen los extremos del cromosoma e impiden que se adhieran a otros cromosomas.
- Bandas: zonas de colores diferentes que aparecen a lo largo del cromosoma cuando son teñidos.
3.1. Tipos de cromosomas.
El centrómero divide a los cromosomas en dos porciones llamadas brazos. La longitud relativa de los brazos permite clasificar a los cromosomas en:
- Metacéntricos: centrómero central y brazos iguales.
- Submetacéntricos: brazos ligeramente desiguales.
- Acrocéntricos: brazos muy desiguales.
- Telocéntricos: centrómero en un extremo. Sólo hay un brazo.
Por otra parte, existen dos tipos especiales de cromosomas enormes llamados cromosomas gigantes:
- Cromosomas politénicos.
- Cromosomas plumosos.
3.2. Números de cromosomas.
Según el número de juegos de cromosomas que posean, las células y los organismos se clasifican en:
- Diploides (2n): células con 2 juegos de cromosomas.
- Haploides (n): células con un único juego de cromosomas.
- Poliploides: células con 3, 4 o más juegos de cromosomas.
4. División celular: mitosis y citocinesis.
La división celular ordinaria consta de dos fases secuenciales: la mitosis y la citocinesis.
4.1.Mitosis.
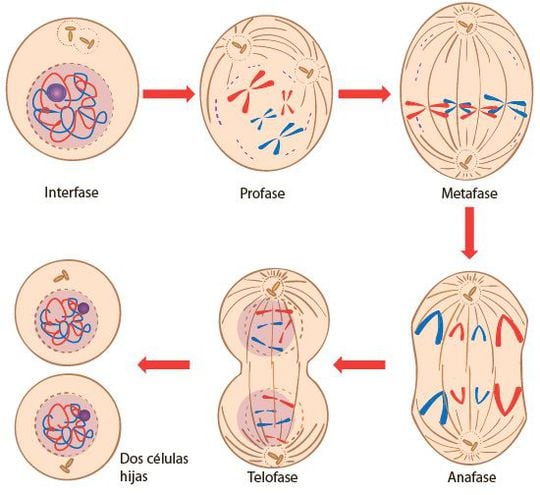
La mitosis es un proceso por el que el núcleo de la célula eucariota se divide en dos núcleos genéticamente idénticos, cada uno conteniendo el mismo número de cromosomas que la célula original.
Aunque la mitosis es un proceso continuo, para sus estudio se divide en 4 etapas: profase, metafase, anafase y telofase.
-Profase.
- El nucleolo se desintegra, ya que el ADN que lo constituye se reparte entre los distintos cromosomas a los que pertenece.
- El ADN en forma de cromatina comienza a condensarse, formándose los cromosomas y haciéndose visibles sus dos cromátidas, unidas por el centrómero.
- Los microtúbulos del citoesqueleto se organizan para formar el huso mitótico, que permitirá el reparto de las cromátidas.
- Al final de la profase se disgrega la lámina fibrosa, la doble membrana nuclear se fragmenta en vesículas y la envoltura nuclear desaparece.
-Metafase.
- La membrana nuclear ha desaparecido por completo.
- Los polares se alargan hacia los cromosomas. Cuando encuentran un cinetocoro cromosómico lo capturan.
- Tras una serie de reajustes, los cromosomas se sitúan en el plano ecuatorial del huso, con un cinetocoro dirigido hacia cada polo. Se forma así la placa ecuatorial, en ella, cada cromátida de cada cromosoma está dirigida hacia un polo de la célula.
-Anafase.
- Anafase A o temprana: la tubulina de los microtúbulos cinetocóricos se despolimerizan, con lo que estos microtúbulos se acortan y arrastran a las cromátidas hacia polos opuestos.
- Anafase B o tardía: los microtúbulos polares, solapados en el ecuador del huso, se deslizan en direcciones opuestas alargándose. Esto alarga la célula y contribuye a la separación de las cromátidas.
- Proteínas motoras, como la dineína citoplasmática y la quinesina, unidas al cinetocoro, se desplazan por los microtúbulos y contribuyen a la separación de las cromátidas.
-Telofase.
- Los microtúbulos polares se alargan al tiempo que los cinetocóricos se acortan hasta desaparecer. Así, las cromátidas llegan a los polos.
- Alrededor de cada grupo de cromátidas se desarrolla la lámina fibrosa y la doble membrana nuclear. El nucleolo reaparece.
- Las cromátidas se descondensan y forman fibras de cromatina.
- Los microtúbulos polares, aún formando parte del huso, se sueldan entre sí y forman un eje que se rompe al tiempo que comienza la citocinesis.
4.2.Citocinesis.
Una vez dividido el núcleo, para completar la división celular debe dividirse el citoplasma y repartir los orgánulos. Este proceso se denomina citocinesis.
La citocinesis suele comenzar en la anafase y acabar entre la telofase y la interfase siguiente.
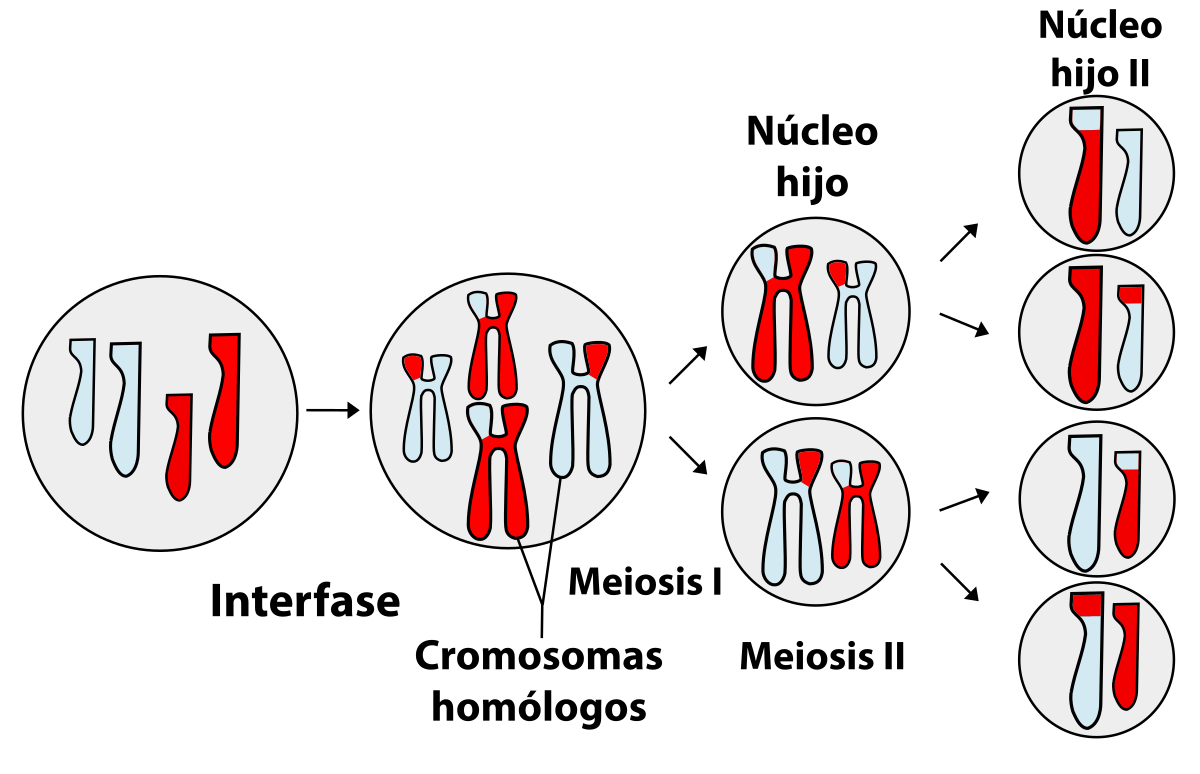
5. Meiosis.
La meiosis es un mecanismo de división celular especial por el que se forman gametos, es decir, células con la mitad de los cromosomas que la célula madre. La meiosis es exclusiva de los organismos con reproducción sexual
La meiosis consta de dos divisiones del núcleo sucesivas.
5.1. Primera división meiótica.
-Profase 1.
Durante ella, la envoltura nuclear y el nucleolo permanecen intactos. Al final, se desintegrarán, al tiempo que se formará el huso acromático. Suele dividirse en 5 fases:
- Leptoteno. Los cromosomas se acortan y ensanchan, haciéndose visibles, aunque no se distinguen las cromátidas hermanas. Los cromosomas están unidos a la lámina fibrosa mediante una estructura llamada placa de unión.
- Zigoteno. Los cromosomas homólogos se aparean y alinean en toda su longitud mediante un proceso llamado sinapsis. La sinapsis ocurre por una estructura proteica llamada complejo sinaptonémico, que une cada gen con su homólogo del cromosoma opuesto. Se forman a sí pares de cromosomas llamados bivalentes.
- Paquiteno. Los cromosomas íntimamente unidos o bivalentes, intercambian fragmentos entre cromátidas no hermanas: sobrecruzamiento. El sobrecruzamiento conlleva una recombinación genética o intercambio de genes, el resultado son cromátidas mixtas, con fragmentos paternos y maternos.
- Diploteno. Desaparecen los complejos sinaptonémicos, por lo que los cromosomas homólogos se separan. Los cromosomas siguen acortándose y condensándose, por lo que se hacen visibles las cromátidas de cada homólogo.
- Diacinesis. Los cromosomas son ya perfectamente visibles como tétradas.Las cromátidas hermanas están unidas por el centrómero. Al final de esta fase desaparece la membrana nuclear y el nucleolo. Los centrosomas se separan y comienzan a formar los microtúbulos del huso acromático.
-Metafase 1.
Se forma una placa ecuatorial doble, con las tétradas en el ecuador celular, que atraviesa los quiasmas de cada tétrada, no los centrómeros.
-Anafase 1.
El acortamiento de los microtúbulos provoca la rotura de los quiasmas. Cada cromosoma homólogo se desplaza hacia un polo de la célula.
-Telofase 1.
El nucleolo y la membrana nuclear son regeneradas. El huso desparece y los cromosomas sufren una ligera descondensación.
-Citocinesis 1.
Se produce la división del citoplasma, originando dos células cuyos núcleos tienen la mitad de cromosomas que la célula madre inicial.
5.2. Segundo división meiótica.
La 2ª división meiótica es muy similar a una mitosis:
- Profase II: muy corta. Desaparecen las membranas celulares y se forman los husos.
- Metafase II: los cromosomas se disponen en la placa ecuatorial.
- Anafase II: se rompen los centrómeros y cada cromátida hermana de cada cromosoma se dirige a uno de los polos.
- Telofase II: se descondensan los cromosomas y se forman de nuevo las membranas nucleares y los nucleolos.
- Citocinesis II: se dividen los citoplasmas. El resultado son 4 células haploide (n) y diferentes entre sí debido a la recombinación genética.
6. Metabolismo celular.
El metabolismo es el conjunto de reacciones encaminado a la obtención de biomoléculas y energía para la actividad vital de la célula.
El conjunto de reacciones que lleva desde una molécula inicial hasta el producto final se denomina ruta o vía metabólica. Algunas rutas son lineales: se parte de una molécula inicial y se obtienen productos finales. Otras son cíclicas: se parte de dos o más moléculas iniciales.
6.1. Catabolismo.
El catabolismo es el conjunto de reacciones degradativas que rompen enlaces de moléculas complejas para obtener otras más sencillas.
En general son reacciones oxidativas y las moléculas que se obtienen se emplean bien como precursores metabólicos para el anabolismo, o bien como carburantes metabólicos, para obtener energía (ATP) y poder reductor.
-Reacciones de óxido-reducción.
Un compuesto se oxida si pierde electrones y se reduce si los acepta. Las oxidaciones biológicas suelen ser reacciones de óxido-reducción, con pérdida simultánea de electrones y protones.
Estas reacciones requieren, por tanto, de dadores de electrones, que suelen ser los carburantes metabólicos; y de aceptores de electrones, que suelen ser los nucleótidos NAD+ que se reduce a NADH + H+ o FAD, que se reduce a FADH2.
6.2. Anabolismo.
El anabolismo es el conjunto de reacciones de síntesis que forman enlaces C-C para obtener moléculas complejas a partir de otras más sencillas.
Este tipo de reacciones requiere energía, poder reductor y metabolitos sencillos llamados precursores.
Anabolismo y catabolismo no son procesos independientes, sino que sus reacciones están acopladas.
7. Catabolismo de los glúcidos.
Los glúcidos son las biomoléculas más utilizadas para obtener energía en los seres vivos. De ellas la principal es la glucosa. Los animales obtienen la glucosa por degradación del glucógeno.
7.1. Glucogenolisis.
Tiene lugar en el citosol. Allí, la enzima glucógeno fosforilasa introduce grupos fosfato en los extremos no reductores del glucógeno, liberando moléculas de glucosa-1-fosfato.
7.2. Fermentación y respiración.
- Fermentación de la glucosa: es una oxidación parcial, en el citosol, en la que el pirúvico no entra en el ciclo de Krebs y los aceptores finales de los electrones y protones del NADH,
- Respiración de la glucosa: es una oxidación total en la que los aceptores de protones y electrones son moléculas inorgánicas.
- La respiración aerobia: se da en la mitocondria. El pirúvico se oxida hasta CO2 mediante la descarboxilación oxidativa y el ciclo de Krebs. El aceptor final de electrones y protones es el oxígeno molecular (O2), que se reduce a H2O.
- La respiración anaerobia: exclusiva de ciertos microorganismos. El pirúvico se oxida a CO2 por rutas diversas. Los aceptores finales de electrones y protones son sustancias inorgánicas.
7.3. Glucólisis o ruta de Embden-Meyerhof.
La glucólisis es una ruta metabólica de 10 reacciones mediante la que una molécula de glucosa es oxidada, mediante fosforilación a nivel de sustrato, hasta formar 2 moléculas de ácido pirúvico, obteniéndose 2 ATP y poder reductor.
- Fase de preparación.
- Fase de oxidación.
- Fase de fosforilación.
7.4. Fermentación de la glucosa.
Las fermentaciones de la glucosa son oxidaciones parciales, sin necesidad de oxígeno, en las que los electrones de la glucosa pasan al NADH y, finalmente, a moléculas orgánicas sencillas.
-Fermentación láctica.
El último aceptor de electrones es el propio ácido pirúvico, que los toma del NADH, dando de nuevo NAD+.
La fermentación láctica es típica de microorganismos de la leche y células musculares.
-Fermentación alcohólica.
El ácido pirúvico se transformará en dióxido de carbono y etanol en dos etapas:
- Descarboxilación del ácido pirúvico: cada pirúvico libera un CO2, originando dos moléculas de acetaldehído.
- Reducción del acetaldehído: los acetaldehído actúan como receptores finales de electrones, reduciéndose a etanol, oxidando al NADH que vuelve a regenerar NAD+.
7.5. Respiración aerobia de la glucosa.
-Descarboxilación oxidativa del ácido pirúvico.
El pirúvico de la glucólisis entra en la matriz de la mitocondria. Allí, gracias a la enzima piruvato deshidrogenasa, pierde CO2 y es oxidado, dando ácido acético, que se transfiere a una molécula de coenzima A para formar acetil-CoA. Los electrones liberados son recogidos por el NAD+, que se reduce a NADH.
-Ciclo de krebs.
- El acetil-CoA transfiere su grupo acetilo a una molécula de ácido oxalacético, dando ácido cítrico y liberando el CoA. Enzima: citrato sintetasa.
- El cítrico pasa a isocítrico a través de un intermediario (ácido cis-aconítico). Enzima: aconitasa.
- El isocitrato sufre descarboxilación oxidativa: pierde CO2 y se oxida (NAD+ -> NADH + H+), dando α-cetoglutárico. Enzima: isocitrato deshidrogenasa.
- El α-cetoglutárico sufre descarboxilación oxidativa: pierde CO2 y se oxida (NAD+ -> NADH + H+). La reacción libera energía, aprovechada para formar un enlace tioéster rico en energía con una molécula de CoA, dando succinil-CoA. Enzima: α-cetoglutarato deshidrogenasa.
- El succinil-CoA se rompe en ácido succínico y CoA. La reacción libera energía suficiente para formar GTP a partir de GDP y ácido fosfórico (fosforilación a nivel de sustrato). Enzima: succinato tioquinasa.
- El succínico se oxida a fumárico (FAD -> FADH2). Enzima: succinato deshidrogenasa.
- El fumárico se hidrata a málico. Enzima: fumarasa.
- El málico sufre una deshidrogenación (NAD+ -> NADH + H+) y se convierte en oxalacético, cerrando el ciclo. Enzima: malato deshidrogenasa.

-Cadena transportadora de electrones o cadena respiratoria.
En la cadena respiratoria, los e- fluyen a favor del potencial de óxido-reducción, desde las coenzimas reducidas NADH y FADH2 hasta el O2. Consta de varios complejos multiproteicos:
- Complejo I (NADH deshidrogenasa): un par de electrones del NADH pasan al coenzima Q (CoQ) mediante el nucleótido FMN y centros Fe-S.
- Complejo II (succinato deshidrogenasa): incluye a la enzima que oxida el succínico en el ciclo de Krebs. Pasa los electrones de la FADH2 al CoQ.
- Complejo III (citocromo b-c1): los electrones pasan del CoQ al citocromo b y de éste a los citocromos c1 y c.
- Complejo IV (citocromo oxidasa): contienen los citocromos a y a3, con iones Cu. Recoge los electrones del citocromo c y, a través de a y a3, los lleva al O2, que se une a 2 protones (H+) de la matriz para dar H2O.
-Fosforilación oxidativa.
La fosforilación oxidativa es un proceso de síntesis de ATP gracias a la fuerza protón-motriz, la entrada de protones en la matriz a favor de gradiente.
-Balance energético del catabolismo de la glucosa.
- Fermentaciones: las únicas moléculas de ATP formadas son las generadas por fosforilación a nivel de sustrato en la glucólisis: 2 ATP.
- Respiración aerobia: en teoría, una molécula de glucosa podría producir hasta 38 moléculas de ATP, sien embargo el balance real sería de 30 o 32 ATP por glucosa.
8. Catabolismo de lípidos.
Los lípidos desempeñan muchas funciones en la célula, una de las cuales es la de suministrar energía, sobre todo a partir de triglicéridos.
Los triglicéridos sufren una lipólisis, una hidrólisis que produce glicerina y ácidos grasos.
9. Catabolismo de proteínas.
Las proteínas sólo se emplean como combustible en casos de ayuno prolongado. En primer lugar sufren proteolisis en los lisosomas o proteosomas, dando aminoácidos.
Los aminoácidos pasan por diferentes rutas metabólicas específicas, en las que pierden el grupo amino y se convierten en pirúvico o en acetil-CoA, entrando en el ciclo de Krebs.
Los grupos amino se eliminarán con la orina como diferentes compuestos nitrogenados.
10. Procesos anabólicos.
- Biosíntesis de aminoácidos y proteínas. Los aa esenciales deben tomarse con la dieta. A partir de estos aa se sintetizan las proteínas.
- Biosíntesis de ácidos nucleicos. ADN y ARN.
- Biosíntesis de ácidos grasos.
- Biosíntesis de glucógeno.
- Gluconeogénesis. síntesis de glucosa a partir de moléculas más sencillas.
10.1. Gluconeogénesis.
Proceso inverso a la glucólisis, salvo algunas reacciones que requieren rutas diferentes y gran aporte de energía.Se parte de 2 moléculas de pirúvico. El pirúvico debe convertirse en 2-fosfoenol-pirúvico. Esta es la etapa más compleja, pues requiere varias reacciones, energía en forma de ATP y GTP y poder reductor como NADH + H+.
11. Anabolismo autótrofo: fotosíntesis.
La fotosíntesis es un proceso anabólico para obtener materia orgánica a partir de la inorgánica y con ayuda de la energía lumínica. Se realiza en cloroplastos de células eucariotas de organismos fotosintéticos.
11.1 Fase lumínica o fotoquímica.
La fase lumínica o fotoquímica consiste en una serie de reacciones fotoquímicas en las membranas de los tilacoides. Se capta energía lumínica y se transforma en energía química.
-Fotosistemas.
- Un complejo antena: son cientos de moléculas de clorofila y otros pigmentos, unidos a proteínas de la membrana, que captan luz de diferente longitud de onda y transfieren su energía al centro reactivo.
- Centro reactivo o centro de reacción fotoquímica: formado por dos moléculas especiales de clorofila dentro de una proteína transmembrana. Actúa como una trampa energética, captando la energía que llega del complejo antena y usándola para impulsar electrones hacia la cadena transportadora de electrones de la membrana tilacoidal.
- Un dador y un aceptor de electrones: diferentes en cada fotosistema.
-Tipos de fotosistemas.
- PS I: se halla por toda la membrana tilacoidal. Contiene dos moléculas de clorofila P700.
- PS II: se encuentra principalmente en los grana. Contiene dos moléculas de clorofila P680, las cuales ceden su electrón a un aceptor primario que es la feofitina.
-Pigmentos fotosintéticos.
- Clorofila: contiene una porfirina unida a un átomo de Mg2+ y a una cadena de fitol, lípido isoprenoide que permite su unión con fosfolípidos y regiones hidrófobas de las proteínas de las membranas tilacoidales.
- Carotenoides: absorben luz verde y azul, protegiendo a las clorofilas de procesos de oxidación.
- Ficobilinas: en ciertas algas y bacterias.Absorben luz verde y amarilla.
-Fosforilación no cíclica.
En esta fotofosforilación los dos PS actúan en serie y los e- siguen un flujo lineal desde el agua hasta el NADP+, que se reduce a NADPH. El flujo electrónico genera energía con la que se produce ATP.
-Hipótesis quimiosmótica de la fotofosforilación.
La fotólisis del agua en el PS II se produce en el estroma. Dos de los e- y 2 de los H+ liberados son aceptados por la plastoquinona (PQ), que se reduce a PQH2.La PQH2 pasa los e- al cit-b6-f y los H+ al espacio tilacoidal. Por tanto, el complejo plastoquinona y cit-b6-f actúa como una bomba de protones hacia el espacio tilacoidal, generando un gradiente electroquímico.Según la hipótesis quimiosmótica de Mitchell, el flujo de protones desde el espacio tilacoidal hasta el estroma tiene lugar a través de la enzima ATP sintetasa, que genera ATP por catálisis rotacional.
-Fosforilación cíclica.
En esta fotofosforilación sólo interviene el PS I y los e- del P700 realizan un recorrido cíclico y vuelven a este mismo centro reactivo. Lo e- pasan a la A0, filoquinona, ferredoxina, plastoquinona y cit-b6-f. Este último libera suficiente energía para crear un gradiente de protones que permite la síntesis de ATP por la ATP sintetasa. Finalmente, los e- del cit-b6-f pasan a la plastocianina, que los devuelve al PS I.
11.2. Fase oscura o biosintética.
Esta fase transcurre en el estroma del cloroplasto y es independiente de la luz.
-El ciclo de Calvin-Benson.
Es un proceso de fijación y reducción del CO2 atmosférico. Es un proceso anabólico y reductor y, por tanto, precisa energía (ATP) y poder reductor (NADPH).
El ciclo de Calvin y la fijación del CO2 por la rubisco son propios de todos los organismos autótrofos. No se dan sólo en fotosintetizadores típicos, como cianobacterias, algas eucarióticas y plantas, que realizan la fotosíntesis oxigénica, sino también en arqueas y bacterias dotadas de metabolismos fotosintetizadores o quimiosintetizadores diversos.
11.3.Fotorrespiración y plantas C4.
La rubisco es una enzima poco eficaz. No sólo por su lentitud catalítica, sino también porque, además de carboxilasa es oxidasa. Por ello, puede unir CO2 a la RuDP para el ciclo de Calvin-Benson, pero también puede añadirle O2 y dar fotorrespiración El que realice una u otra reacción depende de la concentración relativa de CO2 y O2.
No se conoce bien para qué sirve la fotorrespiración, pero parece ser que, en parte, protege a los tilacoides de la fotooxidación que sufren ante un exceso de luz y escasez de CO2.
-Plantas C4: solución a la fotorrespiración.
Las plantas que fijan directamente el CO2 a la RuDP y forman 3PG se denominan plantas C3, por ser de 3 carbonos estas primeras moléculas. Son plantas con gran fotorrespiración.
Sin embargo, existen plantas que fijan primero el CO2 a una molécula de 3 carbonos, dando una molécula de 4 carbonos. Estas plantas se llaman C4 e incluyen a las plantas adaptadas a climas secos y cálidos
11.4. Factores que modulan la fotosíntesis.
- Intensidad lumínica: cada especie tiene un intervalo de luminosidad óptimo. Dentro de ese intervalo, el aumento de la luminosidad aumenta la fotosíntesis hasta un máximo.
- [CO2]: a medida que aumenta, aumenta la actividad fotosintética, hasta llegar a un límite estable.
- [O2]: su incremento disminuye la fotosíntesis debido a la fotorrespiración, sobre todo en plantas C3.
- Fotoperiodo: el rendimiento fotosintético depende de las estaciones, que proveen un número de horas de luz muy variable.
- Temperatura: en general, el aumento de la temperatura incrementa la fotosíntesis, pues aumenta la actividad enzimática. Por encima de cierto valor, la fotosíntesis disminuye. De todas formas, hay plantas adaptadas a climas cálidos y otras a climas fríos.
- Humedad: la falta de humedad provoca el cierre de los estomas y la disminución de la actividad fotosintética. Las plantas C4 son más eficaces en condiciones secas.
- Color de la luz: por encima de una longitud de 680 nm (luz roja) el PS II no actúa, por lo que sólo se da fotofosforilación cíclica.
Fuente: biologeo.
Comentarios
Publicar un comentario
Gracias por comentar. Te rogamos que seas preciso y educado en tus comentarios.