 1. METABOLISMO CELULAR
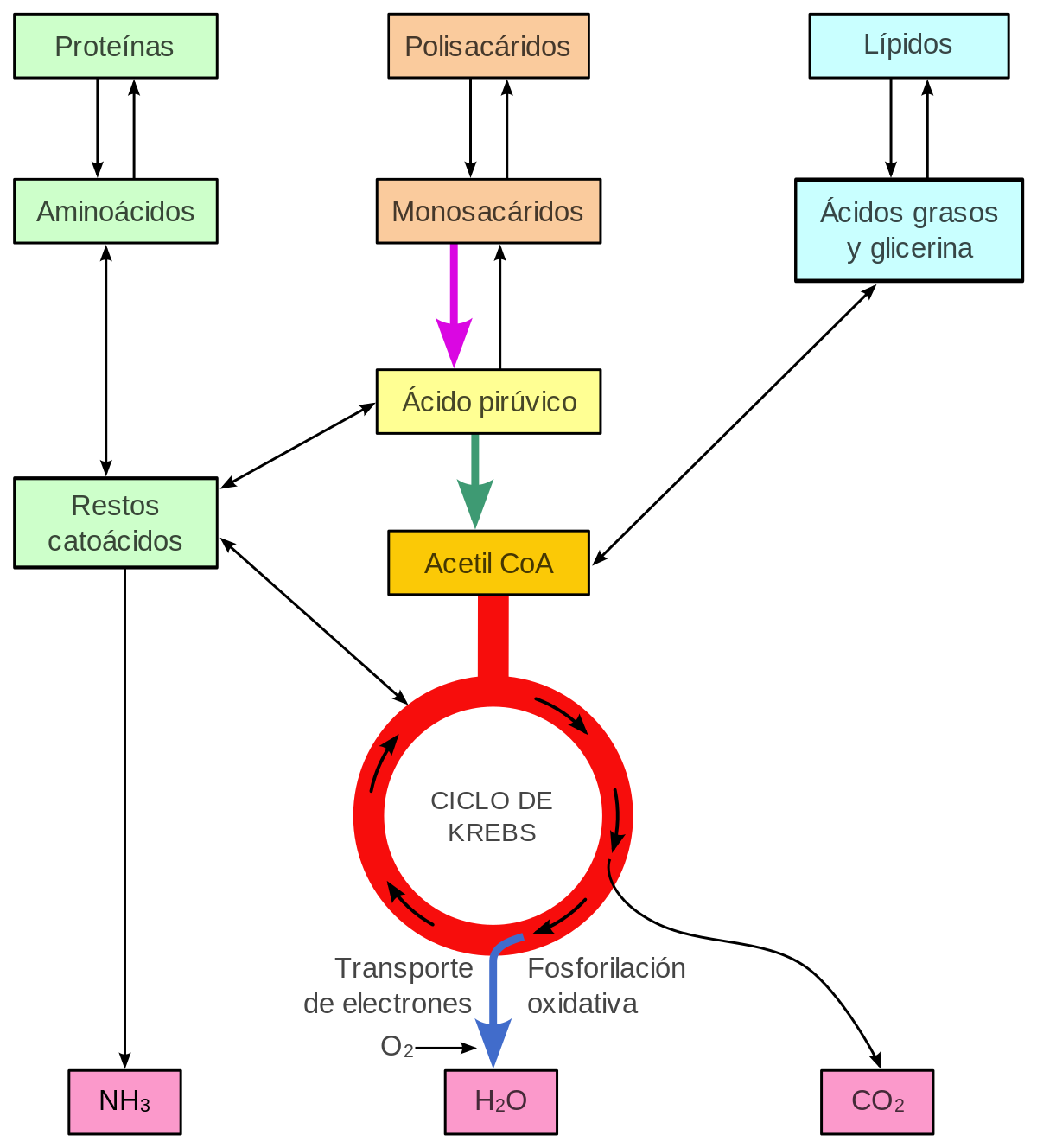
1. METABOLISMO CELULAR Es el conjunto de reacciones encaminado a la obtención de biomoléculas y energía para la actividad vital de la célula. Esto se logra mediante reacciones de biosíntesis que constituyen el anabolismo. La obtención de energía se produce a través de reacciones de degradación de moléculas orgánicas que constituyen el catabolismo.
1.1. CATABOLISMO (DEGRADACIÓN).
El catabolismo es el conjunto de reacciones degradativas que rompen enlaces de moléculas complejas para obtener otras más sencillas.En general son reacciones oxidativas y las moléculas que se obtienen se emplean bien como precursores metabólicos para el anabolismo, o bien como carburantes metabólicos, para obtener energía.
REACCIONES DE ÓXIDO-REDUCCIÓN:
Un compuesto se oxida si pierde electrones y se reduce si los acepta. Las oxidaciones biológicas suelen ser reacciones de óxido-reducción, con pérdida simultánea de electrones y protones. Estas reacciones requieren, por tanto, de dadores de electrones, que suelen ser los carburantes metabólicos.
1.2. ANABOLISMO (SÍNTESIS).
El anabolismo es el conjunto de reacciones de síntesis que forman enlaces C-C para obtener moléculas complejas a partir de otras más sencillas.Este tipo de reacciones requiere energía, poder reductor y metabolitos sencillos llamados precursores.En los organismos autótrofos los precursores son sintetizados a partir de materia inorgánica usando energía lumínicao química. Los heterótrofos obtienen los precursores de reacciones catabólicas sobre compuestos obtenidos en la dieta.
2. Catabolismo en los glúcidos.
Los glúcidos son las biomoléculas más utilizadas para obtener energía en los seres vivos. De ellas la principal es la glucosa. Los animales obtienen la glucosa por degradación del glucógeno del hígado o del de sus reservas citoplasmáticas.
2.1. GLUCOGENOLISIS :
La degradación del glucógeno o glucogenolisis tiene lugar en el citosol. Allí, la enzima glucógeno fosforilasa introduce grupos fosfato en los extremos no reductores del glucógeno, liberando moléculas de glucosa-1-fosfato.
2.2. FERMENTACIÓN Y RESPIRACIÓN:
La glucosa comienza su degradación en el proceso de la glucólisis, en el citoplasma. Se obtienen dos moléculas de ácido pirúvico, ATP y NADH.El destino del pirúvico dependerá del tipo de célula y de la disponibilidad de oxígeno, pudiendo darse:
⦁ Fermentación de la glucosa (es una oxidación parcial, en el citosol)
⦁ Respiración de la glucosa (es una oxidación total en la que los aceptores de protones y electrones son moléculas inorgánicas) lo que distingue a:
-Respiración aerobia: se da en la mitocondria. El pirúvico se oxida hasta CO2 mediante la descarboxilación oxidativa y el ciclo de Krebs.
-Respiración anaerobia: exclusiva de ciertos microorganismos. El pirúvico se oxida a CO2 por rutas diversas.
2.3. GLUCÓLISIS O RUTA DE EMBDEN-MEYERHOF:
La glucólisis es una ruta metabólica mediante la que una molécula de glucosa es oxidada por fosforilación a nivel de sustrato, hasta formar 2 moléculas de ácido pirúvico, obteniéndose 2 ATP y poder reductor (NADH). Se puede dividir en tres fases:
⦁ Fase de preparación: son 5 reacciones en las que la molécula de glucosa se rompe en dos de gliceraldehído-3-fosfato.
⦁ Fase de oxidación: las moléculas de G-3-P son oxidadas por la coenzima NAD+ para formar ácido 1,3 bifosfoglicérico.
⦁ Fase de fosforilación: las 2 moléculas de ácido 1,3 bifosfoglicérico se transforman en ácido pirúvico, obteniendo 4 ATP.
2.4. FERMENTACIÓN DE LA GLUCOSA:
Las fermentaciones de la glucosa son oxidaciones parciales, sin necesidad de oxígeno, en las que los electrones de la glucosa pasan al NADH y, finalmente, a moléculas orgánicas sencillas. Existen numerosos tipos de fermentaciones, destacando:
FERMENTACIÓN LÁCTICA:
El último aceptor de electrones es el propio ácido pirúvico, que los toma del NADH, dando de nuevo NAD+.La fermentación láctica es típica de:
⦁ Microorganismos de la leche: utilizan la lactosa para obtener energía. Es la base de la producción de derivados lácteos.
⦁ Células musculares: cuando el músculo realiza un trabajo muy intenso, el oxígeno aportado es insuficiente para realizar el ciclo de Krebs, por lo que trabaja en condiciones anaerobias, realiza fermentación y produce ácido láctico, dando fatiga muscular.
FERMENTACIÓN ALCOHÓLICA:
El ácido pirúvico se transformará en dióxido de carbono y etanol en dos etapas:
⦁ Descarboxilación del ácido pirúvico: cada pirúvico libera un CO2, originando dos moléculas de acetaldehído.
⦁ Reducción del acetaldehído: los acetaldehído actúan como receptores finales de electrones, reduciéndose a etanol, oxidando al NADH que vuelve a regenerar NAD+.
RESPIRACIÓN AEROBIA DE LA GLUCOSA:
Permite obtener toda la energía de la glucosa gracias a su oxidación mediante una serie de etapas:
DESCARBOXILACIÓN OXIDATIVA DEL ÁCIDO PIRÚVICO:
El pirúvico de la glucólisis entra en la matriz de la mitocondria. Allí, gracias a la enzima piruvato deshidrogenasa, pierde CO2 (descarboxilación) y es oxidado, dando ácido acético. Los electrones liberados son recogidos por el NAD+, que se reduce a NADH.
CICLO DE KREBS:
Ciclo del ácido cítrico o ciclo de los ácidos tricarboxílicos (CAT o TCA). Conjunto de reacciones que oxidan completamente los dos átomos de carbono del acético hasta CO2. El acetil-CoA que inicia el ciclo puede proceder de la oxidación de la glucosa, de los ácidos grasos o de los aminoácidos. El ciclo de Krebs transcurre en la matriz mitocondrial, donde están todas las enzimas necesarias. Sus etapas son:
⦁ El acetil-CoA transfiere su grupo acetilo a una molécula de ácido oxalacético, dando ácido cítrico y liberando el CoA. Enzima: citrato sintetasa.
⦁ El cítrico pasa a isocítrico a través de un intermediario (ácido cis-aconítico). Enzima: aconitasa.
⦁ El isocitrato sufre descarboxilación oxidativa: pierde CO2 y se oxida (NAD+ -> NADH + H+), dando α-cetoglutárico. Enzima: isocitrato deshidrogenasa.
⦁ El α-cetoglutárico sufre descarboxilación oxidativa: pierde CO2 y se oxida (NAD+ -> NADH + H+). La reacción libera energía, aprovechada para formar un enlace tioéster rico en energía con una molécula de CoA, dando succinil-CoA. Enzima: α-cetoglutarato deshidrogenasa.
⦁ El succinil-CoA se rompe en ácido succínico y CoA. La reacción libera energía suficiente para formar GTP a partir de GDP y ácido fosfórico (fosforilación a nivel de sustrato). Enzima: succinato tioquinasa.
⦁ El succínico se oxida a fumárico (FAD -> FADH2). Enzima: succinato deshidrogenasa.7.El fumárico se hidrata a málico. Enzima: fumarasa.8.El málico sufre una deshidrogenación (NAD+ -> NADH + H+) y se convierte en oxalacético, cerrando el ciclo. Enzima: malato deshidrogenasa.
CADENA TRANSPORTADORA DE ELECTRONES O CADENA RESPIRATORIA
Conjunto de reacciones que llevan los electrones desde el NADH y el FADH2 hasta el O2 que, junto a los H+, formará agua.
En la cadena respiratoria, los e- fluyen a favor del potencial de óxido-reducción, desde las coenzimas reducidas NADH y FADH2 hasta el O2. Consta de varios complejos multiproteicos:
⦁ Complejo I (NADH deshidrogenasa): un par de electrones del NADH pasan al coenzima Q (CoQ) mediante el nucleótido FMN y centros Fe-S.
⦁ Complejo II (succinato deshidrogenasa): incluye a la enzima que oxida el succínico en el ciclo de Krebs. Pasa los electrones de la FADH2 al CoQ.
⦁ Complejo III (citocromo b-c1): los electrones pasan del CoQ al citocromo b y de éste a los citocromos c1 y c.
⦁ Complejo IV (citocromo oxidasa): contienen los citocromos a y a3, con iones Cu. Recoge los electrones del citocromo c y, a través de a y a3, los lleva al O2, que se une a 2 protones (H+) de la matriz para dar H2O.
FOSFORILACIÓN OXIDATIVA
Según la hipótesis quimiosmótica, la energía liberada en el transporte de electrones se emplea para bombear protones desde la matriz al espacio intermembranoso.La fosforilación oxidativa es un proceso de síntesis de ATP, la cual la realiza ATP sintetasa, enzima que atraviesa la bicapa lipídica y forma un canal de protones y cuenta con una parte F1, lo que origina un movimiento rotatorio en F1 que produce el ATP: catálisis rotacional.
BALANCE ENERGÉTICO DEL CATABOLISMO DE LA GLUCOSA
⦁ Fermentaciones: las únicas moléculas de ATP formadas son las generadas por fosforilación a nivel de sustrato en la glucólisis: 2 ATP.
⦁ Respiración aerobia: en teoría, una molécula de glucosa podría producir hasta 38 moléculas de ATP.
3.Catabolismo de lípidos.
Los lípidos desempeñan muchas funciones en la célula, una de las cuales es la de suministrar energía, sobre todo a partir de triglicéridos.Los triglicéridos sufren una lipólisis, una hidrólisis que produce glicerina, la cual entrará directamente en la glucólisis.Los ácidos grasos se activarán añadiéndoles CoA, formando un acil graso CoA. Luego, estos sufrirán una ß-oxidación en las mitocondrias o en los peroxisomas. La ß-oxidación de los ácidos grasos dará lugar a moléculas de acetil-CoA, que seguirán la ruta del ciclo de Krebs.
4.Catabolismo de proteínas.
Las proteínas sólo se emplean como combustible. En primer lugar sufren proteolisis en los lisosomas o proteosomas, dando aminoácidos, los cuales pasan por diferentes rutas metabólicas específicas, en las que pierden el grupo amino y se convierten en pirúvico o en acetil-CoA, entrando en el ciclo de Krebs.
5.Procesos anabólicos.
La fotosíntesis es un proceso anabólico para obtener materia orgánica a partir de la inorgánica y con ayuda de la energía lumínica. Se realiza en cloroplastos de células eucariotas de organismos fotosintéticos. La fotosíntesis consta de 2 fases:
5.1. FASE LUMÍNICA O FOTOQUÍMICA
La fase lumínica o fotoquímica consiste en una serie de reacciones fotoquímicas en las membranas de los tilacoides. Se capta energía lumínica y se transforma en energía química.La incidencia de fotones sobre los fotosistemas libera electrones que reducen el NADP+ a NADPH y permiten la síntesis de ATP.
FOTOSISTEMAS:
Son la unidad estructural de la membrana tilacoidal. Están formados por:
⦁ Un complejo antena (cientos de moléculas de clorofila y otros pigmentos), unidos a proteínas de la membrana, que captan luz de diferente longitud de onda y transfieren su energía al centro reactivo.
⦁ Centro reactivo o centro de reacción fotoquímica: formado por dos moléculas especiales de clorofila dentro de una proteína transmembrana. Actúa como una trampa energética, captando la energía que llega del complejo antena y usándola para impulsar electrones hacia la cadena transportadora de electrones de la membrana tilacoidal.
⦁ Un dador y un aceptor de electrones: diferentes en cada fotosistema.
TIPOS DE FOTOSISTEMAS:
Los vegetales superiores presentan en sus membranas tilacoidales dos tipos de fotosistemas:
PS I: se halla por toda la membrana tilacoidal. Contiene dos moléculas de clorofila P700. La clorofila P700 cede sus electrones a un aceptor primario que es una molécula llamada clorofila A0. El hueco electrónico se llena con un electrón procedente del dador, la plastocianina, que se encuentra en la cadena transportadora de electrones.
PS II: se encuentra principalmente en los grana. Contiene dos moléculas de clorofila P680, las cuales ceden su electrón a un aceptor primario que es la feofitina (Feo). El hueco se rellena con un electrón procedente del dador, el agua. Por tanto, en este fotosistema sucede la fotolisis del agua, en la que una molécula de agua se rompe dando dos electrones, dos H+ y O2 que se desprende.
PIGMENTOS FOTOSINTÉTICOS
Todos los organismos fotosintéticos tienen varios tipos de pigmentos para captar la energía lumínica. Entre ellos:
⦁ Clorofila
⦁ Carotenoides (carotenos y xantofilas)
⦁ Ficobilinas: en ciertas algas y bacterias.
FOTOFOSFORILACIÓN NO CÍCLICA (OXIGÉNICA)
En esta fotofosforilación los dos PS actúan en serie y los e- siguen un flujo lineal desde el agua hasta el NADP+, que se reduce a NADPH. El flujo electrónico genera energía con la que se produce ATP. La representación de este proceso se denomina esquema Z:
⦁ Los fotones que llegan al PS II son recolectados por su complejo antena que libera un e- de alta energía. Este e- pasa por la cadena de transporte. Los e- llegan finalmente al PS I. Durante este transporte se libera energía que se emplea en producir ATP a partir de ADP + Pi: fotofosforilación.
⦁ Los fotones que llegan al PS I son captados por el P700, que libera un e- de alta energía, que será transportado por otra cadena. Finalmente, acaba en el NADP que se reduce a NADPH. Los huecos electrónicos del PS I son rellenados por los e- que llegan del PS II.
HIPÓTESIS QUIMIOSMÓTICA DE LA FOTOFOSFORILACIÓN
⦁ La fotólisis del agua en el PS II se produce en el estroma. Dos de los e- y 2 de los H+ liberados son aceptados por la plastoquinona (PQ).
⦁ La PQH2 pasa los e- al cit-b6-f y los H+ al espacio tilacoidal. Por tanto, el complejo plastoquinona y cit-b6-f actúa como una bomba de protones hacia el espacio tilacoidal, generando un gradiente electroquímico.
⦁ Según la hipótesis quimiosmótica, el flujo de protones desde el espacio tilacoidal hasta el estroma tiene lugar a través de la enzima ATP sintetasa, que genera ATP por catálisis rotacional.
FOTOFOSFORILACIÓN CÍCLICA (ANOXIGÉNICA)
En esta fotofosforilación sólo interviene el PS I y los e- del P700 realizan un recorrido cíclico y vuelven a este mismo centro reactivo. Lo e- pasan a la A0 cit-b6-f, el cual libera suficiente energía para crear un gradiente de protones que permite la síntesis de ATP por la ATP sintetasa. Finalmente, los e- del cit-b6-f pasan a la plastocianina, que los devuelve al PS I.Es por esto que en este proceso se obtiene ATP, pero no O2 ni NADPH.
6.2. FASE OSCURA O BIOSINTÉTICA
Esta fase transcurre en el estroma del cloroplasto y es independiente de la luz. Se emplea el ATP y el poder reductor (NADPH) obtenidos en la fase luminosa para reducir moléculas sencillas oxidadas. Así se consiguen moléculas orgánicas sencillas que se utilizarán en otros procesos.
EL CICLO DE CALVIN-BENSON
Es un proceso de fijación y reducción del CO2 atmosférico. Es un proceso anabólico y reductor y, por tanto, precisa ATP y poder reductor.Esta reacción está catalizada por la enzima ribulosa-1,5 difosfato carboxilasa oxigenasa (Rubisco). Se necesita en grandes cantidades.El ciclo de Calvin y la fijación del CO2 por la rubisco son propios de todos los organismos autótrofos.Seguidamente, el 3GP se reduce a gliceraldehído,mediante ATP y NADPH.
El G-3-P puede ahora seguir dos rutas:
⦁ 10 de cada 12 moléculas de G-3-P seguirán el ciclo de Calvin
⦁ 2 moléculas de G-3-P pasan al citosol, donde son precursoras de la síntesis de glucosa (gluconeogénesis).
6.4. FACTORES QUE MODULAN LA FOTOSÍNTESIS
 El rendimiento de la fotosíntesis depende de numerosos factores ambientales, entre ellos:
El rendimiento de la fotosíntesis depende de numerosos factores ambientales, entre ellos:⦁ Intensidad lumínica
⦁ [CO2]
⦁ [O2]
⦁ Fotoperiodo
⦁ Temperatura
⦁ Humedad
⦁ Color de la luz
Fuentes: BIOGEO
Comentarios
Publicar un comentario
Gracias por comentar. Te rogamos que seas preciso y educado en tus comentarios.